Genetics
[Translated to English from Swedish by Eva Gunnarsson, Nikopeja's Siberians, and Ulrika Olsson. Proofreading by Riëtte van Beek, Nidoba Maine Coons]
By Ulrika Olsson
- Introduction
- Basic genetics
- Different types of inheritence
- The color genetics of cats
- Cat genetics for coat, ears and tail
- The inheritence of some diseases and defects
- Breed to avoid genetic diseases
- How do we fight already existing health problems?
- Summary
Why is one supposed to learn all this? After all, it is a hobby, not a profession!
In order to be able to breed in a good way one needs to know some genetics. And yes, cat breeding is "only a hobby", but that doesn't change the fact that genetic knowledge is needed, hobby or not. The breeds and with that the cats can be harmed, both in short and long term, if breeding is done in an inappropriate way. It is of course not ethically justifiable to harm cats just because of lack of knowledge. So if one doesn't have the time or interest in learning this, then one shouldn't breed cats. You can always show cats, and have fun with your cats in other ways!
Here you will get some basic knowledge on the subject. But we do recommend you to learn more on your own, by reading the recommended literature and other books related to the subject, or by going to lectures when an opportunity arises.
Basic Genetics
The cell
All organisms consist of cells. They live either as singular cells or as larger or smaller aggregations. Bacteria and microscopic creatures consist of one single cell, but higher plants and animals consist of many thousands, millions or billions of cells. In such complicated organisms the cells have different tasks and are gathered in organs with different functions. Examples of such organs are for instance brain, kidneys, and lungs or in plants leaves and pistils.
Form and size of a cell can vary very much. All of them can be seen though as a mixture of biological molecules inside a bag. The "wrapping" is called cell membrane and the "soup" within, the cytoplasm, contains a large amount of easily recognized structures, which all have a specific function. The largest structure is the cell core, called "nucleus", which contains the chromosomes.
The chromosomes are paired, so that if for example a gene for eye colour is on one chromosome in a pair then there is also a gene for eye colour on the same location, same "locus", on the other chromosome. It is not necessary the same predisposition for a certain eye color on both chromosomes, but it might be. Genes with similar functions are called homologous.
When a cell divides itself a doubling of the number of chromosomes takes place in that each and every of the chromosomes splits itself in half along the middle into a group of two. The pair is kept together in a certain place called the centromere. Since each cell has two copies of each chromosome to start with, this means that every cell, just before division, has 4 copies of each chromosome. The two original chromosomes of the pair are similar, but the two copies of each chromosome that are held together at the centromere, are identical! After this doubling, the centromeres line up in the center of the cell, where the centromeres split: the two chromosomes of the identical pair, now finally released from each other, moves to opposite sides of the cell. Since all chromosomes do this, the chromosomes are divided into two identical groups, each group moving to opposite sides of the cell to form the nucleus of the daughter cell. The cell wall is constructed between the two daughter cells and the process can start all over again. This type of cell division is called mitosis. Thus, during the mitosis the mother cell is divides into two identical daughter cells, which both have exactly the same set of chromosome as the mother cell.
Mitosis:
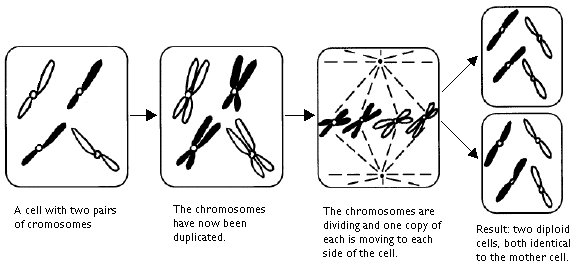
There is also another type of cell division: reduction division or meiosis. During meiosis the numbers of chromosomes are reduced so that the daughter cells only contain half as many chromosomes as the mother cell, one chromosome from each pair. These cells with a single set of chromosomes are said to be haploid (Greek: haploos=single). Cells which have sets of double chromosomes, that is all chromosomes in pairs, are called diploid (Greek: diploos=double). In higher organisms only gametes, reproductive cells like eggs and sperms, are haploid. If the gametes were not haploid the number of chromosomes would double itself in each generation.
Meiosis:
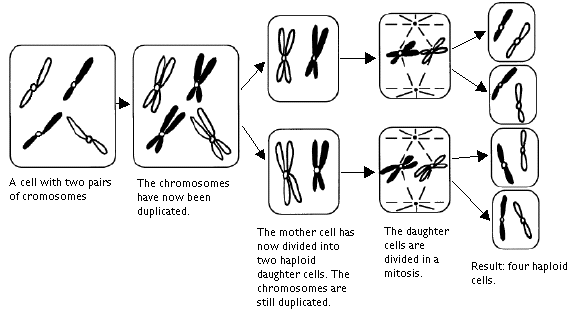
In several organisms, including mammals, there are two of the male chromosomes that do not form a homologous pair. They are called the x- and y-chromosomes. During the meiosis, two types of sperm are always formed, one with an x-chromosome and one with a y-chromosome. The female cells contain two x-chromosomes and each egg contains one of these. When an egg and sperms melt together during conception the chromosomes are mixed together and the diploid order is restored. One chromosome of each pair of chromosomes is hence from the mother and the other one from the father. The fertilized egg starts growing by mitosis. The sex of the new organism depends on whether the egg has been fertilized by a sperm with an x-chromosome, which gives a female, or a y-chromosome, which gives a male. Among certain species, birds for example, it is the other way around in that it is the egg cell that has two different gender deciding chromosomes.
Genes
The word gene, first introduced in 1910, was used as an abstract hereditary unit controlling a specific hereditary characteristic within a specific species. The existence of the genes was predicted in studying hereditary characteristic, like for instance the color of flowers, from known "parents" for many generations. The most famous studies were done by Mendel, an Austrian monk, on various characteristics of pea plants. The factor found deciding the color of flowers was present in different versions. In one case it gave white flowers, in another red. The same thing applied to the structure of the seed surface: some are wrinkled, others are smooth. These different varieties of one specific gene are called alleles. Each organism has two alleles for each characteristic, one from each parent. In every generation they are split up when the gametes are created during the reduction division (meiosis): each haploid germ cell has only one allele of the original pair. At conception a new combination is created. Both the alleles in the pair can be identical, and the individual is then said to be homozygotic (Greek: homos=same, zygon=pair) for that pair of alleles. If we call the alleles deciding the color of the pea flower for fr (red color) and fw (white color), an individual can have the following combinations: frfr = homozygote, fwfw = homozygote, frfw = heterozygote. The germ cell will have either the allele fr or fw.
The alleles for different hereditary characteristics is often transmitted to the gametes independently of each other, since the different genes often are placed on different chromosomes or far from each other on the same chromosome. A pea plant can for instance have the alleles frfw color of the flower and slsk for long respectively short stem. The gametes can then contain either of the following combinations: frsl, frss, fwsl or fwss. When the gametes are formed the alleles are transmitted for color, fr and fw, independently of the alleles for the length of the stem, sl and ss.
Crossover
Sometimes the genes for two different characteristics are placed on the same chromosome. If the gene for flower color and the gene for stem length would be on the same chromosome, one might not expect to get both red flowering and white flowering pea plants with both long and short stems. Instead, one would expect the allele for long stem to follow the allele for red flowers, so that one would only be able to get red flowering high plants or white flowering small plants. Well, now this is not the case, for during the reduction division the two paired homolog chromosomes cross over each other and exchange parts. This is called crossover or recombination. As a result, the chromosome that an individual gets from its mother contains parts of the chromosome from the mothers mother, and parts of the same chromosome of the mothers father. Though, still it is so that alleles placed on the same chromosome tend to follow each other. The closer they are the less is the likelihood that a crossover will take place in between them, and the greater the chance that the alleles will follow each other. The genes are said to be linked.
Mutations
A chromosome is a very, very long DNA molecule. Each genes consist of a little piece of such a DNA-molecule. Sometimes something goes wrong during the duplication of the chromosomes so that a part of the DNA-molecule, maybe part of a gene, is different from the original molecule. The cell with this incorrect DNA-molecule then divides, and the fault is spread further. Even gametes can have an altered DNA-molecule and then all cells in the offspring, or new organism, will have the same defect. This alteration is called a mutation. Some of such mutations don't affect the individual at all. Others may have such a drastic consequence that the individual will not be able to survive, but some mutations will give altered but healthy individuals. Example of such mutations could be the gene for non-agouti and dilution in cats. Some mutations even have a positive influence in the survival of the individual compared to other. Then this new predisposition will spread and animal and plant species have been improved through mutation. This is the basis of the theory of evolution. The natural selection will take care that only animals who are fit will survive, while unhealthy mutations will be weeded out.
Different types of inheritence
Dominant - recessive
Gregor Johann Mendel, the father of genetics, experimented with pea plants. As we mentioned before, one of the things he looked at (that he studied) was the colour of the flowers. He started with two pea species which in cultivating had shown to be stable with regards to the colour of the flower. One strain gave always red flowers and the other strain only white flowers. Mendel crossed these strains by transferring pollen from one kind to the pistil of the other. The result was plants which had only red flowers. These plants then were allowed to reproduce by themselves. This time the offspring was not homogenous. About three quarters had red flowers and one quarter of the plants had white flowers. One calls this a splitting between two different types with a splitting ratio of 3:1, which shows the number plants with red flowers and those with white flowers (see below). Mendel assumed that all plants, which resulted from that crossing between the two types, contained the predispositions for both read and white. He assumed also that the red color had a stronger influence than the white. He hence called the stronger influential predisposition for dominant. The weaker capacity he called recessive. He declared that the dominant gene for red flowers completely hid the recessive gene for white flowers in the heterozygotic plants.
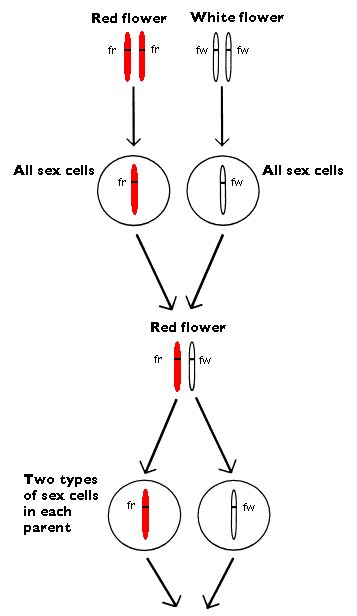
There can be two different types of gametes present in each of the parents:
Gametes from the male - fr and fw
Gametes from the female - fr and fw

Incomplete dominance
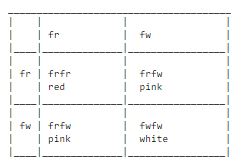
The dominance of one allele over an other is not always complete. Sometimes one can distinguish a homozygotic individual from a heterozygotic one. One clear example of this one can find in the Four O-Clock flower, Mirabilis. Here the red color is incomplete dominant over the white. Heterozygotic flowers get at color in between red and white - they become pink. This is called incomplete dominance.
If one crosses a red and a white Mirabilis all the offspring will have pink flowers. If one crosses two pink flowers we get the following breeding chart:
Over-dominance
Sometimes the phenotype for the heterozygotic individual is more extreme than both the homozygotic varieties. That is, if pp gives a light pink flower and PP gives a dark pink flower, then Pp gives a dark red flower! And that feels a little unexpected. This phenomenon is called over-dominance and occurs among other things in the immune system: individuals that are heterozygotes for genes of the immune system, are often healthier and have a better chance of survival than individuals that are homozygotic (with whatever allele) for these genes.
Polygenes
Sometimes two or more genes can contribute to one specific characteristic. This has been studied for example on wheat. Wheat seed has as a rule a red hue, but even pure white seed exist. When crossing between a stable red seed variety and a white one, one only gets red seed. In the next generation the white seeds surface again. This depends on that wheat have two different genes for the color of the seeds, which are placed in one chromosome each (inherited hence independently of each other). The first crossing can be written as R1R1 R2R2 (red) x r1r1 r2r2 (white), which the gives R1r1 R2r2 (red). The dominance by R1 and R2 is incomplete, and due to this the effect of the genes can be added. This results in that the red colored wheat seeds can have different hues of red. Crossing between two of the genotype R1r1 R2r2 will give the following crossing table and colors:
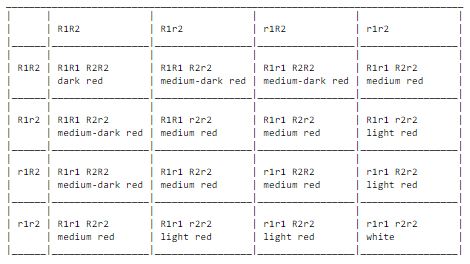
We then get a scale of five different nuances while only having two pair of genes involved. When it comes to for example the size of a cat, length of fur or the amount of undercoat, there certainly are many more genes involved. The result will be a continuous scale between two extremes.
Sex-linked heredity
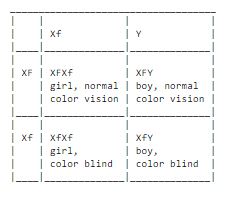
Color blindness in humans is an example of a predisposition that is a sex linked inheritance. The predisposition is recessive and placed on the x-chromosomes. To mark this we can call it Xf, and the dominant gene for normal color viewing we can call XF. It is commonly known that there are a significant amount of men who are color blind than there are color blind women. The reason for this is that a color blind woman needs to have the defect on both her x-chromosomes, otherwise the dominant gene for normal color vision would override so that she didn't show the defect. A man on the other hand only has one x-chromosome and if the gene for color blindness is situated on it then he is color blind. There is no gene for color vision on the y-chromosome that could correct the defect.
The consequence of this will then that for a woman to be color blind she must have inherited the defective gene from both her father and from her mother. The father is then color blind, since the gene cannot be hidden with men. The mother on the other hand doesn't have to be color blind; she can carry it as a hidden gene.
A color blind man must have inherited the disposition from his mother, since he has have inherited the y-chromosome from his father and hence the x-chromosome must come from his mother, and with the x-chromosome comes the defective gene.
For example: we make a mating scheme between a color blind man and a woman who carries a hidden predisposition:
Epistasis
Some alleles are not only dominant over other alleles belonging to the same locus but they also obscure the effects of other genes. Or maybe it is recessive towards another allele on the some locus, but still obscuring the effects of any allele in another locus. This is called epistasis. One example of this is the gene for white in cats, W. This gene is epistatic to all other genes for coat color - whether the cat carries genes for black, blue, red, agouti or spotted, it will only be the white color that is showing.
The color genetics of cats
When starting as a breeder one should learn how colors are inherited in cats. First of all one need to know what the different colors and patterns look like. At least one should know the colors of the breed one plans to breed. Different colors can be studied at shows - ask the cat owners if you are unsure what the color of a cat is, or to see if you had it right. Maybe your cat club can arrange a seminar in color classification?
When you have learned the different colors it is time to learn about color genetics.
The original domesticated cat is a shorthaired brown agouti cat. All other colors and varieties that exist have arisen later as mutations from this.
Red or black pigment
Cats as you know have many different colors, but there are only two different types of pigments: eumelanin, black, and phaeomelanin, red. A male cat can only have one type of pigment, while a female cat can have both (tortoiseshell and tortie). This is because the gene for pigment type is situated on the x-chromosome. The inheritance is thus sex-linked. A male has one x-chromosome and one y-chromosome, and hence, can only have EITHER the allele for black pigment or the allele for red pigment. Females on the other hand have two x-chromosomes and the can have BOTH an allele for black and an allele for red. None of the alleles dominates over the other so the cat will be both red and black - tortie. A female can of course have the allele for black on both x-chromosomes. She will then be black. She can also have the allele for red pigment on both chromosomes, and then she will be red.
Denomination: Xr = x-chromosome with an allele for phaeomelanin, red pigment
Xb = x-chromosome with an allele for eumelanin, black pigment
Y = y-chromosome, no gene for pigment
Alternatively: O = red (orange)
o = black
Remember that a male always must inherit the y-chromosome from his father and thus his x-chromosome will come from his mother. A male cat's pigment will always come from his mother, while his father's pigment is not inherited. A female cat on the other hand gets her x-chromosome from her father. She inherits thus her father's pigment. And she also gets one of her mother's x-chromosomes. She thus inherits pigments from both her mother and her father. Notice that this is only true for genes on the x-chromosomes, so for the colors of the cat it is only true for red versus black. Silver, agouti/non-agouti, dilution and other is inherited regardless of sex.
For example: A tortie female is mated to a black male. The female can either give Xb or Xr to the kittens. The male can give Xb or Y.

The chance of getting a black female is one to four, 25 %. The chance of getting a black male is also one to four, 25%, exactly as the chance in getting a tortie female and a red male.
Agouti or non-agouti
In an agouti cat some of the hairs are striped, while others are fully colored. In a brown agouti cat, for example, the hairs in the black part are black, while the brown areas are mostly black- and brownish striped hairs.
The non-agouti variety corresponding to brown agouti is black. The allele for non-agouti is recessive. Both parents must thus carry the non-agouti allele for an offspring to become non-agouti.
The non-agouti gene doesn't work that well on red cats. Non-agouti red cats show the same tabby markings that they would show if they did have the agouti-allele. One usually can decide whether a red cat is agouti or non-agouti by looking at the area between the nose and the mouth. An agouti cat is white or very light colored in that area, while a non-agouti cat has approximately the same color there as in the rest of the face. The same thing applies to cream colored cats. If the cat is red-white or cream-white and has a white spot over the nose it will of course be impossible to decide whether the cat is agouti or non-agouti by using this method. One can look at the rim of the ears - they are lighter on an agouti than on a non-agouti.
Denomination: A = agouti
a = non-agouti
Patterns
Agouti cats can be blotched (or classic) tabby, mackerel tabby, spotted, or ticked (Abyssinian).
A mackerel tabby is, like the name suggests, a cat with vertical relatively small stripes on the body. A classic or blotched tabby has broader bands and the pattern is more marbled. Spotted means that the cat has dots and a ticked cat has a dark stripe along it's back continuing out on the tail tip while the rest of the cat is pure agouti without any markings. Abyssinians and Somali are example o breeds that have ticking, and this pattern is often called Abyssinian.
The ticked tabby pattern is inherited with a dominant allele. The recessive allele is for non-ticked.
Denominations: T = Ticked tabby, Aby pattern
t = not ticked tabby
If the cat is not ticked tabby, it can have the classic/blotched tabby pattern or the mackerel tabby pattern. Mackerel is dominant over classic/blotched.
Denominations: Mc = mackerel tabby
mc = classic/blotched tabby
Note that all cats carry genes for patterns, even if they are non-agouti so that the pattern doesn't show. Also, a ticked tabby cat carries gene for classic/mackerel even if it doesn't show. This is because the ticked tabby pattern is epistatic over all the other patterns, meaning that ticked is sort of "dominant" over the genes for other patterns. And non-agouti is epistatic over all patterns, including ticked. It is hiding all the patterns.
One doesn't know the exact genetics behind a cat becoming spotted. There might be one or more genes breaking up the bands on a tabby or the stripes of a mackerel into spots. It is generally considered that the spotted cats that have tabby as a foundation will have the sharpest dots. It is not unusual for cats to have both stripes and dots, so it might be hard to decide whether a cat is mackerel or spotted.
The gene for dilution
The dilution gene causes a modification so that the color becomes lighter. This is because the pigment granules are forming clumps. To the eye the coat looks lighter, in the same way as a mixture of black and white grains of sand look grey. The allele for dilution is recessive. It makes a black cat blue and a red cat cream. A brown tabby cat becomes a blue tabby.
Denominations: D = non-dilution, black/red
d = dilution, blue/cream
The genes for chocolate and cinnamon/sorrel
The allele for chocolate is causing the hairs to become lighter for other reasons than the allele for dilution. In chocolate cats the black pigment granules, which normally are round, are a bit flattened. This enables more light to be reflected - the coat looks paler. This allele is also recessive. A cat which otherwise would have been black becomes, as the name implies, chocolate colored. A blue cat becomes lilac. The chocolate allele doesn't work on the red pigment, so red stays red and cream will be cream. The chocolate allele first came from the Siamese and can therefore be seen in all breeds that have the Siamese pointed colors recognized. This is also present in the Oriental Shorthairs, which are very closely related to the Siamese.
The cinnamon/sorrel allele flattens the pigment granules even more than the gene for chocolate which makes the coats seem even lighter. This allele is recessive to both the allele for chocolate and the allele for normal pigmentation. An oriental with this color is said to be cinnamon colored and it describes the color perfectly. The sorrel color of the Abyssinian is genetically identical to the cinnamon of the Oriental, but since the Abyssinian is ticked while the Oriental is non-agouti they don't look that similar. Earlier the sorrel colored Abyssinians were called red, but the breeders back then realized rather quickly that they couldn't be genetically red since there were never born any tortoiseshell colored Abyssinians.
A diluted cat with cinnamon/ sorrel genes in homozygotic alleles is called fawn. This cat can be said to be more beige or ivory colored. This gene does not work on red and cream.
Denominations: B = normal pigment, black
b = chocolate (brown)
bl = cinnamon/sorrel (light brown)
The albino series
(Himalayan/Siamese coloring, Burmese coloring, albino)
The albino series consists of five alleles. The gene for normal color, for example black, is dominant and is represented by C. cb is the gene that gives the paler color of the Burmese, the black cat becomes brown. cs gives Siamese coloring, ca gives a white cat with blue eyes, a blue eyed albino. This cat is not to be mixed up with the more commonly dominant white cat, which also can have blue eyes. And at last there is a gene called c. It gives genuine albino cats, which are white cats with red eyes.
The cb allele is not fully dominant over the cs allele. A cat with the genotype cbcs has a coloring inbetween Burmese color and Siamese masking. The color of the eyes is turquoise. This coloring represents a special breed, not yet recognized in FIFe. It is called Tonkinese and cannot be pure bred. In a mating between two Tonkinese there is a 25% chance of getting Siamese masked kittens, 50% chance of getting Tonkinese and 25% of getting Burmese colored kittens.
The Siamese colored kittens are born completely white. The color of the mask doesn't appear until after a week. This is due to that the cells of Siamese colored cat cannot develop any pigment if the temperature is too high. When the kittens are in the womb of their mother's they are warm and comfortable, and no pigment at all is developed. When they are born it becomes cool enough for the pigment to develop on the cooler extremities, like for instance ears, legs and tail. This temperature dependant pigment production is called acromelanism. Acromelanism is present in other animals, for instance in rabbits.
The color snow leopard, present in Bengals, is primarily a masked cat with relatively weak contrasts between mask and body. The dots can therefore be seen all over the body, but the markings are a bit darker in the face, on the legs and on the tail.
Foreign white is a Siamese with the gene for dominant white, W (see below). The result is a completely white cat with the dark blue eyes of the Siamese. A normal white blue eyed cat has considerably lighter eyes.
Denominations: C = normal colour
cb = Burmese
cs = Siamese
ca = blue-eyed albino
c = true (red-eyed) albino
Silver
The silver gene is dominant, and it inhibits the development of pigment in the coat. The under coat will be white, while the tip of the hairs will keep its colors. In an agouti cat the most obvious effect can be seen in the lighter areas, between the dark tabby markings. A silver cat that is non-agouti is called smoke.
Denominations: I = silver (inhibitor of melanin)
i = non-silver
Dominant white and white spotting
The gene which makes a cat pure white is dominant. This allele inhibits the normal function of the coloring cells, so that no color is produced. A completely white cat thus carries the genes for red/black, agouti/non-agouti, dilution/non-dilution, etc. The gene for white is thus epistatic.
Pure white cats often have a color spot on the head when they are born. It disappears when the cat has developed its adult fur. This spot can reveal what color the cat has underneath the white.
White cats can have yellow eyes, blue eyes or one blue and one yellow eye (odd-eyed). One doesn't know how the eye color in this case is inherited.
White cats are often deaf. This is due to a degenerative change of the cochlea in the inner ear. It is definitely more common with deafness in blue-eyed white cats than in yellow-eyed, but there are yellow-eyed white cats that are deaf and blue-eyed cats with normal hearing. Cats can also be deaf on only one ear. Odd-eyed cats are often deaf on the ear which is situated on the same side as the blue eye.
The main gene for white spotting is situated in the same locus as the gene for solid white. This allele seems to be incomplete dominant over the allele for non-white, meaning that a cat with two alleles for white spotting gets more white than a cat with just one. There is an overlap though - bicolour cats might be either homozygous for the white spotting allele or heterozygous. Since we have this variation is expression, there must be more genes that modify the expressions of these genes, since there are not three sharply divided groups of cats with white spotting, but instead the variation seem to be continuous.
The allele for all white is however dominant over the allele for white spotting.
Denominations: W = white
ws = white spotting w = non-white
So a cat with genotype wsws would be anything from bicolour to van, while a cat with wsw can be anything from fairly low grade white to bicolour. But a cat with the genotype Wws looks just the same as a cat with genotype Ww - it is all white either way.
Breeders of Birman can hardly be satisfied with the above mentioned theory. Birmans have white paws, and when two Birmans are mated all kittens get white paws. One never gets any cat completely without white and one never gets a harlequin or van colored cat.
Hence one has come up with the theory of a recessive gene for white paws. This is sometimes denominated as g (gloves). GG is according to this theory a cat without white. Gg could possibly give a cat with a white medallion or a white spot on the tummy and gg would give white paws and maybe a bit of white on tummy and throat (these spots would be concealed in the light coat of the Birman). A gene has been found that is thought to be responsible for this, but it doesn't seem to quite fit with the reallity, so better ignore that for now.
The Chinchilla gene
Earlier one believed that chinchilla and shaded silver were only extreme varieties of silver agouti. Now many believe that there is a special gene responsible for these color variations (polygenes are also involved). The assumption is that this gene will cause to widen the light bands of the agouti hairs, which causes the whole hair, except the outermost tip, to become light. Even the single colored hairs on an agouti cat will be light. The gene doesn't work on non-agouti cats. The gene is called Wb (widening of band) and is incompletely dominant. WbWb gives shaded silver or shaded golden and wbwb gives a normal agouti coat. Cats with the genotype A- WbWb are also said to be tipped.
On newly born tipped and shaded kittens one can almost always see signs of tabby/mackerel/spotted. The kittens will get lighter for a long period before they get their final coloring.
Rufism
Rufism is a term representing the relatively large variation of brown pigmentation in cats. The amount of rufism decides whether a brown tabby cat will have a warm brownish color or a more grayish color. It will also decide whether a red cat will be a warm dark red or a pale orange. The variation between these two extremes is continuous. Hence, this characteristic is probably influenced by many other genes (polygenetically inherited), where many genes add small positive or negative effects.
Some consider the brown spots that one sometimes sees on silver cats, also as highly graded rufism, while others think that this is not connected to rufism.
The amount of rufism is often very important when breeding for color, as one does with Persians. In brown agouti cats one wishes to have as much rufism as possible. And a wild colored Abyssinian without a high grade of rufism wouldn't look particularly impressive. In red cats, one gets a Êdeep warm red color only with a lot of rufism: the more rufism, the warmer the color. A silver tabby show Persian should not have any brown spots. If you breed silver cats and support the theory that the brown in silver cats are caUsed by rufism, you will want as little rufism as possible. Even other colors are affected by rufism, but not as much as in the above mentioned variations.
Extra reading on tortoiseshell cats
What makes some areas in a tortoiseshell red and others black?A few days after conception, when the original cell has divided itself in many similar cells, one of the x-chromosomes is shut down in each cell. For the rest of the life there is only one x-chromosome in the cell that is functioning. Randomly it's decided which x-chromosome is to shut down. When the cells again are dividing themselves the daughter cells will have the same x-chromosome shut downed as the mother cell. This will make the cat have a mix of red and black markings.
Tortoiseshell males
A male can't be tortoiseshell since he only has one x-chromosome. Still, sometimes a tortoiseshell male appears in a litter. How can this be? Must he then not have two x-chromosomes? Yes, this is exactly how some of the events of the tortoiseshell males are explained. Some of them have something wrong with their chromosomes, they have two x-chromosomes and one y-chromosome. Either the mother put two x-chromosomes in the egg, or the father gave both his x-chromosome and his y-chromosome to the sperm. The same chromosomal aberration occurs in many other animals and in humans. It is called Klinefelter's syndrome. This chromosome aberration leads to sterility - when a male tortoiseshell is not sterile, then he is tortoiseshell because of another reason.
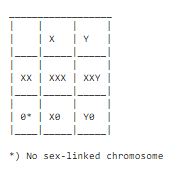
XXX - meta-female, to humans: look normal, at least some of them, maybe all, are fertile.
XXY - Klinefelter's syndrome.
X0 - Turner's syndrome, to humans: short in growth, childish, often sterile. Mice with Turner's syndrome look like and appear just like normal mice.
Y0 - dies early in development of the foetus.
The tortie coloring of a male may also be caused by a fault in the pigmentation of the skin. This is often seen in a red cat, that has some spots, sometimes even large spots, of black. These males breed as a red male. In principle, one might get a black tomcat with a bit of red. Such a cat however has never been reported.
Yet another variety of tortoiseshell males may be caused when two fertilized eggs, maybe already grown to a group of cells, are united and becomes one embryo. This is the reverse of one fertilized egg splitting into two embryos, that become identical twins. If one of the fertilized eggs would have become a black tomcat, while the other would have become a red tomcat, then the combined embryo will be a tortie male cat. There has for example been born a red-blue and white tabby Maine Coon in the US, who has been shown to be the result of such a combination of fertilized cells. The male is fertile and has given only the red color to his kittens.
Of course, these anomalies can also occur where both X-chromosomes carry the same color. Then the result is not a tortie tom, but a plain normal red (cream) or black (or variant) male cat. These males will never be suspected of being something special.
If you are interested in seing pictures and reading more details, you can look at this private website.
Cat genetics for coat, ears and tail
Longhair and shorthair
The gene for longhair is recessive. The differences in length of coat one can see in comparing a Persian with a semi-longhair cat, for instance a Ragdoll, is due to modifying polygenes, that is genes with plus or minus effects, depend on the variation of the degree of longhair to shorthair.
Denominations: L = shorthair
l = longhair
Cornish Rex - r
Cornish Rex has awn hairs and down hairs, but have no guard hairs. The coat hairs are shorter than normal and wavy. The whiskers are also shorter than normal and often bent. This rex gene, which is recessive, appeared in 1950 in Cornwall, England. The curly coat of the German Rex seems to be caused by this gene too.
Devon Rex - re
The Devon Rex has a coat similar to Cornish Rex, but at a closer inspection one can see that the Devon Rex - as opposed to the Cornish Rex, has guard hairs. The coat hairs of the Devon Rex are even shorter and more extreme than the Cornish Rex. This gene is recessive and has another locus than the r-gene. If one mates a purebred Cornish Rex - rr ReRe - with a purebred Devon Rex - RR rere - all kittens will have normal coats (shorthair) with the genotype Rr Rere. The Devon rex-gene appeared in Devonshire, England in 1960.
Hairlessness
As well as there are several different genes giving curly coats there have also surfaced different mutations causing hairlessness in cats. One was discovered in France in 1930s. It was called h and was recessive. The most known mutation appeared in Canada in the beginning of 1970. This gene has been taken care of in the breed of the Sphynx. The gene causing the hairlessness of the Sphynx is also recessive and has the denomination hr. There are only these two variations that have been examined by genetics, but hairless cats have been found in different parts of the world, like for instance Australia, Mexico and Morocco.
Manx
The Manx is the well-known tailless cat, said to originate from the Isle of Man in Great Britain. The breed is relatively old, but also controversial, since some defects may follow the Manx gene. The gene is incomplete dominant and lethal in its homozygotic form. In heterozygotic form it might, apart from giving a very short tail or taillessness, give more or less shortened vertebrae, anomalies in the pelvic region with problems in movement as consequence, neurological problems and distorted intestines. There are however different opinions on how severe these problems are. The gene is denominated M.
Japanese Bobtail
Japanese bobtail is a very old breed from Japan. It has approximately one decimeter long tail which is often bent or curled and stiff. This type of tail is caused by a recessive gene. Unlike the Manx-gene this gene doesn't seem to affect the cat in any other way.
American Curl
The breed American Curl is founded on a gene which gives a defect on the external ear, the ear is bent backwards. The effect of the ears is developed when the kitten is between twelve and sixteen weeks old. The first cat of this kind first appeared in 1981 in California. The gene, denominated Cu, is dominant. It seems like this gene is of no harm to the cat, whether it is in heterozygotic or in homozygotic form.
The inheritence of some diseases and defects
Hypertrophic Cardiomyopathy - HCM
Hypertrophic cardiomyopathy, or HCM, is the most common heart disease among cats. The disease, which also exists in humans, causes enlarged heart walls in the affected individual. The disease cannot be seen from birth, it evolves gradually. Some cats die from the disease as kittens, but the most common is that the cat is free of symptoms for many years. At an ultrasound most of the cases, but not all, be found at the age of three if the examining veterinarian has good equipment and is knowledgeable in the field, but an affected cat can in some cases live long with its disease. Often the cat has no symptoms until it suddenly dies of a serious arrhythmia, or by a blood clot often situated in the back legs. The cat can also develop heart insufficiency. Possible medical treatment is only meant to relieve the symptoms, it is not a cure.
The disease appears in Maine Coon and British Shorthair to be inherited mainly as a dominant gene. Other breeds have not been studied in regard to this, but there is reason to believe that the inheritance is similar. The development of the disease varies however between different breeds, hence one suspect that there might be several different mutations causing the disease within different families. This is also the case in humans.
Polycystic kidney disease - PKD
Cats, who are afflicted by polycystic kidney disease, or PKD, will in time get more and larger cysts in the kidneys. These will force away the normal kidney tissue, so that the function of the kidneys is growing more reduced. Little by little the cat will feel ill, it will loose weight, doesn't want to eat. It also drinks more than usual. One can make it easier on the cat's kidneys by giving it diet food for kidney problems, but there is no cure for the disease itself.
The disease is inherited by single dominance. Experienced veterinarians with good equipment find most cases, but not all, at the age of one year.
Hip Dysplasia - HD
When one hears about hip dysplasia most people think of dogs, but the problem exists also among cats. Hip dysplasia is a hereditary defect in the pelvic capsule making it not as deep as it normally is. This causes the hip capsule not exactly fit into the pelvic capsule and the surfaces start starts to grind against each other. This might cause pain when the cat is moving. Cats are very good at hiding pain and can suffer from their HD even though they are not limping. Instead they might move a little bit calmer or less and avoids high jumps. Cats with mild HD don't have to have any complications.
HD in cats is believed to be inherited polygenetic recessive, which means that two HD-free cats together can have offspring with HD. Two cats with HD might have HD-free kittens, but the chances for normal hips are of course better if the parents have good hips.
Patella luxation - PL
Patella luxation is a defect where the kneecap (patella) has a tendency to slip out of its position, it luxates. The cause of this might be that the trochlear groove in which the kneecap rests has to shallow and rounded sides and/or that the ligaments keeping the kneecap in place are too loose. PL can also be caused by an accident. The symptoms are that the cat suddenly stops when walking, with the leg stretched out to the rear. By backing a little the cat might get the kneecap to jump back into the right position. The cat might also be hurting in the knee and getting inflammations. Cats with symptoms can be operated.
PL seems to be polygenetic.
Progressive retinal atrophy - PRA
This disease causes the retina to degenerate. In time the cat will be completely blind. There is no cure for the disease, but if the cat lives indoors, and if the furniture is not moved around to much in the home, even blind cats manage quite well and can live a long life.
PRA exists in different varieties, one is hereditary dominant, one recessively, and then there is one variety that one doesn't know if it's hereditary.
Gangliosidosis - GM1 and GM2
Gangliosidosis is a hereditary disease (actually two, GM1 and GM2) caused by a defective enzyme. This results in that a substance is stored in the brain of the affected cat. The first symptoms can be seen at the age of 2-4 months, when the cat starts showing bad coordination in the back legs, weakness and tremor. This gradually gets worse and leads to paralysis, blindness and epileptic seizures. The disease is lethal.
There are, as mentioned above, two variations of this disease, GM1 and GM2. Both are caused by single recessive genes. Fortunately a DNA-test exists for both varieties, so one can test relatives to the afflicted cats with a blood test.
Umbilical hernia
An umbilical hernia is a hole in the white ligament, where the abdominal muscles are jointed. Through this hole, if one is unlucky, parts of the intestines might fall out. This can cause a blockage in the intestines so that they burst. The result might be a life threatening inflammation in the abdomen. Umbilical hernia should be operated in order to prevent this.
The hereditary of umbilical hernia is unclear. It might be different types of inheritance in different families and breeds. Umbilical hernia might also be caused by accidents.
Flat chest
Kittens with this defect look as a rule normal at birth, but at approximately 1-2 weeks of age the chest falls inward and becomes compressed. Even the back might at the same time become bent. When this happens the kitten will gain little weight, maybe even the weight falls a bit, but then it picks up in a fairly normal rate, if the defect is not too substantial. In slight cases the flatness might reverse completely before the kitten is ready to move to a new home, so that it will not even be a remark on the veterinarian's examination. Of course such a kitten shouldn't be used in breeding, and the buyer should be informed of that the kitten has had a flat chest. In severe cases of flat chest it might be less room for the internal organs as the kittens grow, so that it will not be able to survive. Between these two extremes are the cats that easily get tired and out of breath, but in spite of this can live with the defect.
The hereditary has been studied on Burmese and it seems to be a single recessive gene in that breed.
Chryptorchidism
Sometimes it happens that a male kitten's testes, or one of them, don't descend from the abdomen to the scrotum. Instead they, or it, will stay in the abdomen or in the groin. This is when a cat is called chryptorchid. Testes that are not in place will not be able to produce sperms, but they will produce sexhormones. A bilateral chryptorchid cat will act just as a normal male, unless he is neutered, even though he is sterile. There are studies proposing a higher risk of cancer in testis's that are malplaced, and this might be yet another reason to neuter chryptorchid males, even if this means searching for the wandering testis in the abdomen, which sometimes isn't that easy.
One doesn't know how chryptorchidism in inherited, but it is for sure SOME type of "sex-restricted" inheritance. That the inheritance is "sex-restricted" means that even females can carry genes for chryptorchidism, even if they of course don't show it themselves. If one for instance wants to continue with a kitten in a litter where there is a chryptorchid kitten, it is safer to choose a normal male than choosing a female. The female might have the same genotype for chryptorchidism as the brother who has the defect and passing on the problem to her sons in the same way as the chryptorchid male would do, while a normal brother shows that he has a better genotype in regards to this.
Hereditary muscle dystrophy (myopathy, spasticity)
There is a hereditary disease, usually just called myopathy (earlier "spasticity"), which causes degeneration of the muscles in shoulders and pelvic. The first symptoms of the disease usually show sometime between the ages of 2 weeks and 6 months. The afflicted kitten usually is drawn back and more quiet than its siblings. They then get a problem with keeping the head up, and they may also walk a bit stilted. The disease, which has no cure, will lead to death, usually before the kitten has reached adulthood.
The disease is inherited by a single recessive gene.
Kinked tails
Kinked tails show themselves now and again on all cats in all breeds, even in domestic shorthair. It is a defect on the tail, very often on the outermost tip of the tail, which can vary a bit in type and in degree. Some kinked tails can be seen already on the newborn kitten, but they can also take as long as 1? years before it is shown. Before that the skeleton is not fully outgrown, and hence hereditary tail kinks might gradually evolve.
It is not known how kinked tails are inherited, but a single recessive gene or polygenes have been suggested. Probably there are different types of kinked tails, which also are inherited differently. A kinked tail might also be caused by an accident.
Breed To Avoid Genetic Diseases
The risk of genetic diseases is something that is discussed more and more often among cat breeders and in the different cat clubs and associations. Of course we all want our cats to be healthy! Still bad genetic diseases show up every now and then in different breeds. Why does this happen? What can we do about it? In order to understand what is happening in our breeds when the genetic "epidemics" show up, and in trying to control them, one needs to have a little knowledge about the basics of population genetics.
An important word: Gene frequency
Using the gene for dilute color as an example, let us assume that we have a breed population of 100 cats. Since every cat has a double set of chromosomes, this population will have 200 loci for the dilution gene - meaning 200 places where the D or d gene can be situated. Now assume that 40 of these loci are filled with a d-gene, while the remaining 160 are filled with D-genes. Then the gene frequency for d in this population is 40/200 = 0.20 = 20%. In the same way we find that the gene frequency for D is 160/200 = 0.80 = 80%.
Effective Population
The fact that some breeds are larger in population than others and also the fact that the gene pool can be too limited in a small breed is probably obvious to most breeders. But far too many breeders feel secure in that the larger breeds - for instance Persians and Birmans - have a large enough gene pool. No risk of inbreeding problems there, unless you choose to deliberately inbreed a line! But that is not always true.
We sometimes discuss the fact that overbreeding from a few outstanding individuals is harmful for breeds, even the larger breeds. The most extreme overbreeding from one individual that we could imagine is if one single male is mating every female in a population in a generation. With that kind of breeding, of course the gene pool will not be large, even if the population contained a thousand unrelated females. In order to get a better idea about just HOW large the gene pool really is in such a case, we could calculate the effective population. If the population contains 100 individuals, with an equal number of males and females, all of which are mated randomly to each other with the same amount of offsprings resulting from every parent, then the EFFECTIVE population is also 100.
On the other hand, with the extreme overbreeding of one male described above, we can calculate the effective population with the following formula:
1 1 1
---- = ------ + ------
Ne 4 x Nm 4 x Nf
Where Ne = the effective population, Nm = number of males, Nf = number of females.
Generally speaking, the effective population will not be larger than 4 times the number of individuals of the least represented sex. (Unless you have set up a breeding program specificly designed to avoid losing genetic variation, but that is very seldom the case in cat breeding.) This means that if 5 males are used, the effective population will not be more than 4 x 5 = 20, even if we could use a million different females in that breeding program.
In reality of course it is seldom the case that five males are used for equally many litters and the remaining males in the population are not used for breeding AT ALL. Then it gets a bit difficult to use this formula. But don't worry! There are other methods!
A too small effective population will cause the degree of inbreeding to increase for every generation. There is a connection between inbreeding and effective population. Using this connection we will be able to calculate the effective population of our cat breeds. We can use our pedigrees to calculate the coefficient of inbreeding, COI. The easiest way of doing this is to enter the pedigree in a good pedigree program that is capable of calculating the COI. It is also possible to calculate it by hand. It is not difficult at all, but if the relationships are complicated and you want to calculate the COI from many generations, it is very time consuming, and there is a considerable risk to make a small error somewhere along the way. However, if the relationships are simple and the number of generations to calculate from is reasonable, you could quickly calculate the COI directly from the pedigree.
Simple calculation of the inbreeding coefficient
Even though there are computer programs, which will calculate the inbreeding coefficient automatically, based on pedigrees entered into a database, it is still handy to be able to calculate the inbreeding in less complicated pedigrees manually. Here is a short description on how to calculate the inbreeding coefficient from a pedigree by using the path coefficient method.
The Path Coefficient Method
Start drawing an "arrow pedigree", where every individual only appears once. The arrows should point from parents to offspring and always point downwards, or diagonally downwards.
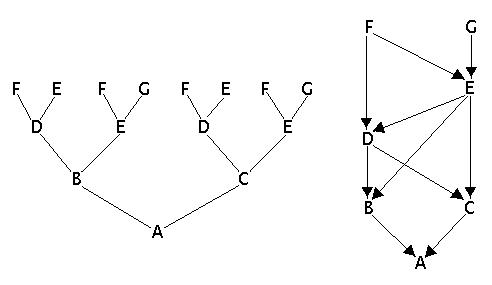
Example 1
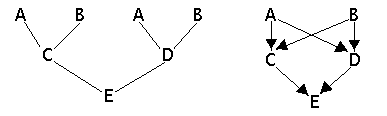
Example 1: Ordinary pedigree and arrow pedigree.
In the arrow pedigree we search all paths that go from one parent to the other without passing any individual more than once. For each path we count the number of individuals involved. The contribution from each path to the inbreeding coefficient is 1/2 raised to the number of individuals involved in the path.
From the arrow pedigree in example 1 we then get two paths from one parent to the other:
CAD -> (1/2)³ = 1 / (2 x 2 x 2) = 1/8
CBD -> (1/2)³ = 1 / (2 x 2 x 2) = 1/8
The inbreeding = 1/8 + 1/8 = 1/4 = 0.25 = 25%
If the common ancestor, i.e. where the path turns and goes downwards again, is inbred we have to take that into account. (For the two paths in the example above the common ancestors, A and B respectively, are underlined.) Calculate the inbreeding coefficient for the inbred common ancestor by using the path coefficient method. Add 1 to this inbreeding coefficient and multiply with the contribution given by the path in question.
So if the number of individuals along a path is n, and the inbreeding coefficient for the common ancestor is F, the total contribution to the inbreeding coefficient will be (1/2)^n x (1+F).
The sum of the contributions of all the paths will then be the inbreeding coefficient.
Example 2
Example 2: Ordinary pedigree and arrow pedigree.
The existing paths are BDC, BDFEC, BEFDC, BDEC, BEDC, BEC.
The common ancestor D is inbred and the arrow pedigree for him looks like this:
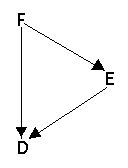
Arrow pedigree for D
The inbreeding in D is then (1/2)² = 1/4.
The contribution to the inbreeding coefficient from each path will then be:
| Path | n | F | (1/2)^n x (1+F) | total |
|---|---|---|---|---|
| BDC | 3 | 1/4 | (1/2)³ x (1+1/4) = 1/8 x 5/4 = 5/32 | = 0.15625 |
| BDFEC | 5 | 0 | (1/2)^5 x (1+0) = 1/32 | = 0.03125 |
| BEFDC | 5 | 0 | (1/2)^5 x (1+0) = 1/32 | = 0.03125 |
| BDEC | 4 | 0 | (1/2)^4 x (1+0) = 1/16 | = 0.0625 |
| BEDC | 4 | 0 | (1/2)^4 x (1+0) = 1/16 | = 0.0625 |
| BEC | 3 | 0 | (1/2)³ x (1+0) = 1/8 | = 0.125 |
The inbreeding = the sum of all contributions = 0.46875 = 46.875%
What happens if the effective population is too small?
One thing that happens is that the inbreeding coefficient increases for every generation. Actually, this happens for ALL populations that are not of unlimited size, but then the natural selection is probably working against more inbred individuals, so that reasonably small increases of the inbreeding are adjusted and status quo is kept. It is also known that a larger number of eggcells get fertilized than the number of offsprings that finally are born in the litters, and one theory is that these early featuses have to "fight" for their place in the uterus, and that the more homozygous featuses are less likely to survive. This theory however not been proven to be true.
What will happen, when the inbreeding coefficient is increased generation by generation? In the beginning, not much at all. It isn't until the degree of homozygosity reaches a certain critical level that the real problems show up, and then it is usually much more difficult to take corrective action. It is much better to start working against these problems before the symptoms show up. The pedagogic problem then of course is that those who begin to breed with too few individuals won't immediately see the problems that it will cause. They think "I have bred in this way for many years, and I have not had any problems". But as we can see, "trial and error" is not a very good approach here! When "error" appears it is a bit late to adjust in an easy way.
Why then is inbreeding so dangerous? One thing - that every educated breeder knows about - is that it means an increased risk of doubling up on harmful or lethal recessive genes. The double set of chromosomes otherwise protects us from this risk, to a great extent in a population that is not too inbred. All individuals carry a few harmful recessive genes. Some people think that the inbreeding clears out the harmful recessives and leads to a healthier breed for the future. But first, the inbreeding doesn't clear out anything in itself, it has to be combined with a strong selection in order to clear out any undesirable genes. Second, you have to inbreed incredibly strongly in order to get all or almost all loci homozygous, so that you can see what the cats are carrying and weed out all the undesirable genes. Mate a female with her full brother, and 25% of all loci are homozygous. Then mate two of these offsprings with eachother, and 37.5% of their loci are homozygous. And then we take two of THOSE offsprings and mate them with each other! Now the inbreeding is so strong that most breeders would back away. But still "only" 50% of the loci are homozygous. So in spite of this drastic inbreeding, we will miss out on exposing other recessive, potentially harmful genes.
But let us assume that we go all the way with this! We breed a line towards 100% homozygosity, selecting strongly against harmful genes all the way. All individuals will then have exactly the same genotype, except for the fact that the males must have a Y chromosome where the females have a second X chromosome.
OK, it took a lot of effort and money to make this so called isogene line, and many cats died on the way. But if we now finally have reached to this point, then we have a line that is 100% healthy from a genetic point of view! Yippeeee!!!
It is possible, this can be done, if you are careful not to let the level of homozygosity increase more quickly than you manage to weed out the bad genes. This has been done with mice that are to be used for scientific tests. It works very well! But... they only manage to get about one line out of twenty to survive. The other 19 lines are dying in the process. Maybe better not to take that chance?
Also, the immune system is not very good in homozygous individuals. The immune system works much better if the loci involved are heterozygous, since this gives the individual the possibility to develop more DIFFERENT kinds of antibodies, not just lots of antibodies of the SAME kind. This is not a major problem in laboratory mice, since their environment is quite protected from (undesired) contagious diseases, and since it isn't exactly considered to be a tragedy, unfortunately, if a laboratory mouse dies. If on the other hand a much loved pet cat and family member dies it is indeed very sad. Hmmm... Perhaps not such a good idea after all?!
On top of that mutations happen spontaniously and would with time destroy our fine genotype. You have to count on one or two new mutations in every individual.
I think we had better change our strategy!
But what if the breed is already inbred?
If a breed or a population is already so inbred that clear signs of inbreeding depression have shown up, for instance a high rate of early cancer or infections, what to do then?
If there are unrelated lines in other countries, of course the best solution would be to increase the exchange of cats between those countries. If such unrelated lines are not available, we will have to outcross to another breed or unregistered cats that fit the standard reasonably well. If enough new genes are mixed into the population, the inbreeding problem will be solved.
A not uncommon objection against these kinds of solutions is that we don't know which new harmful recessives might be introduced into our breed through these outcrosses. That is true, we don't know. What we do know though is that most individuals carry some harmful recessive genes. Many breeders also think that it is better to have a more inbred population less different kinds of genetic diseases, in order to more easily keep them under control. Maybe there are even tests available for those deseases. But, as we will see below, it is better to have lower frequencies of several different harmful recessives than to have a higher frequency of one single recessive.
Assume that we have a population A with a gene frequency of 50% for some kind of recessive defect. We will compare it to a population B with gene frequencies of 10% for five different recessive defects. Both populations will then have the same frequency of harmful genes, but population A has defect genes of only one kind (easy to keep under control) while population B has its defect genes divided in five different kinds.
The risk for a kitten in population A to show the genetic disease is then 0.50 x 0.50 = 0.25 = 25%.
The risk for a kitten to show a genetic disease in population B is 5 x (0.10 x 0.10) = 0.05 = 5%.
This shows that we get considerably less defective kittens in a population which has lower frequencies for several different kinds of diseases. The most effective way to keep a breed healthy is not to try to eliminate the harmful recessives, but to get the frequency down to a such a low level that two harmful recessives of the same kind almost never meet.
Some breeders will hesitate to outcross because they are afraid that the type will be gone for ever. Some breeders are of the opinion that inbreeding (linebreeding) is the only way to get excellent and uniform type. It is true that by using inbreeding you can achieve quicker results in this area. The problem is that you risk the long term health of the cats. It is possible to achieve the same result without inbreeding, although it takes more time. Unfortunately inbreeding is a very tempting shortcut for breeders that are interested in showing their cats. But one should keep in mind that most of the genes that are doubled up by inbreeding have absolutely nothing to do with type. For instance, a human has approximately 30 000 genes, and 98,5% of those are identical with those of a chimpanzee! And still, aren't we rather different from a chimpanzee? How large a part of the genes could different between a Siamese and a Persian? Or a Norwegian Forestcat and a Maine Coon? Or between a Burmese of good type and a Burmese look alike moggie with reasonably good type? Not more than what we can fix with some generations of selective breeding, I'm quite sure about that!
Genetic problems are showing up in the breeds!
Yes, that is also due to a too small effective population! Unless it is due to a breeding without regard to the anatomic functions of the animal. Breeding for extremely long bodies might cause problems with the back, and breeding for very short faces might give problems with the teeth, breeding for extremely triangular, square, round, etc. heads might cause problems with jaws, eyes, brain, or whatever. A cat must be allowed to first and foremost be a CAT. It is not a piece of clay that we can shape after our own estetic ideals. A cat doesn't consist of circles, triangles, squares, or other geometric figures, we have to remember that. Maybe we should breed all cats with the coats of a poodle, so that we could CUT out the geometric shapes and odd angles that we find attractive. Then the cats could have their anatomy in peace. No, even if the standard says that a head should be triangular or square, we as breeders must resist to go to extremes. It should be the head of a cat - not a geometric figure.
Except for this breeding for extremes, it is the too small effective populations that cause the high frequencies of many genetic diseases to show up in breeds. Many breeders seem to be a bit confused about this. They might think that if we have for instance 10% of the cats in a breed affected by PRA, meaning a gene frequency of approximately 32% for the recessive PRA gene, and if we don't test and work to reduce this frequency, then the frequency will automaticly increase with time. This is of course not correct. If it were, then also the frequency of diluted (blue, cream, etc.) cats would increase all the time unless we selected against the dilution gene. If the effective population is big enough, and no selection for or against PRA is made, then the gene frequency will stay at 32%.
On the other hand if we select ever so weakly against PRA, for instance let the cats who themselves have PRA (homozygots) have no more than one litter, then the gene frequency will decrease. Slowly with a weak selection, more quickly with a strong selection.
But then what happens if the effective population is too small? What will then happen with the gene frequency? It will be the same effect as if you toss up a coin 10 times. Your chance for heads is 50% every time. And if you had tossed up this coin 1000 times, you would have got quite close to 50% heads and 50% tails. But now you only toss it 10 times. Then it is not very surprising if you by chance got 70% heads and 30% tails, or 30% heads and 70% tails, or something like that.
In the analogous scenario in a small cat population, it means that the gene frequency of around 30% in the next generation might have increased to 35%, because of this random effect. Or else it might have decreased to 25%, because of the same random effect, which in the PRA case would of course be a lot nicer. But let's be pessimistic and assume that the frequency increased to 35%. Then the EXPECTED value of the gene frequency for the next generation is also 35%. But by chance it might end up at 29%, 34%, 38%, 42%, or whatever. The smaller the effective population is the larger the risk is of getting a large deviation from the expected value of the gene frequency. Then this frequency, that we got randomly, will be the expected value for the next generation. This phenomenon is called random drift. If the impact of this random drift gets stronger than the impact of the selection - natural or artificial - then the changes of the gene frequency could very well be the opposite to what we wanted. DESPITE the selection. Then the eyes might get paler in our Siameses, or the lynx tufts might get smaller in our Norwegian Forestcats, or PKD might get more common in our Persians. That would of course be anything but funny!
If we now look into why PKD became a far too common problem in the Persians, it could hardly be caused by some mysterious selection in favour of lumpy kidneys. It must have another cause.
It must of course have started with a mutation in a cat long ago. It was a dominant gene, so the cat developed cycts on its kidneys. Let us assume that it was a male that died from the PKD at 5 years of age. Or perhaps at 7-8 years of age. Either way, we have a certain selection against the gene. If the population is then large enough, the frequency will then decrease and eventually go down to 0%. And even if there were no selection against the gene what so ever, there would be a good chance for the gene to disappear within a few generations, since the frequency could by chance be slightly larger or slightly smaller. And since the frequency was initially very small (one mutated gene in a large population), it is rather likely that the frequency by chance happened to decrease to 0%, and then the gene is gone.
So, the effective population for the Persians apparently was not large enough. Random drift resulted, and by chance this unfortunately caused an increase of the frequency of the PKD gene. In spite of a certain amount of selection against the gene, the result was that the frequency ended up at approximately 25-30% before more breeders became aware of the problem and a stronger selection was introduced.
What does all this tell us? That if we don't have large enough effective populations, then high frequencies of unpleasant genetic problems will continue to pop up. If we are unlucky we might also have difficulties attempting to reduce these problems with selection.
If we instead make sure that we have large enough effective populations in our breeds, genetic diseases will not pop up as a common problem in the entire population. And as a bonus we avoid inbreeding depressions and bad immune systems.
To breed with too small effective populations, and at the same time start projects to fight against genetic diseases within a breed, is just like being treated for lung cancer and continue smoking. Or to scoop out and wipe away the water that have poured over the rim of the bath-tub, while we still haven't remembered to close the tap, so the water is still pouring INTO the tub.
To work for large enough genepools is a kind of preventive medicine measures for the breeds. It doesn't seem clever just to treat the ailments, without bothering about the preventive measures that could prevent the ailments to arise in the first place.
We should also keep this need for reasonably large effective populations in mind when we create and accept one spotted shorthaired cat breed after the other, and one large furry semilonghair breed of medium head-type after the other, etc. Unless the number of breeders willing to work with spotted shorthaired breeds is increased at the same rate as the increase of the number of breeds, the reqruitment of for instance Bengal breeders - just an example - will be at the expense of the Ocicats', Spotted Oriental Shorthairs', Egyptian Maus', etc. possibilities to keep a constant and large enough number of animals for breeding in their breeding programs. Can these breeds afford this? Will the new breed be able to create a place for itself among all these other breeds? Or perhaps they will all get too small populations, so that we in the end have destroyed all the spotted shorthair breeds? These are important things for us in the cat organisations to think about. These threats are real, not just "in theory", and we have already begun to see the first effects, although not yet as badly as in the dog breeds. However, now we have the chance to avoid getting into as bad problems as the dog breeders! We can learn from their mistakes and change into a healthier way of breeding, or we can continue as before and end up in problems.
So this is something that we have to start working with. It must not be forgotten among all the specific projects against specific diseases. It is about the actual basis for the health of the cats and breeds.
How do we fight already existing health problems?
Problems in single cats
If a cat is diagnosed with a defect or a disease, which might be hereditary or not, how does one act? One basic rule is to not breed with that cat, but to continue with its relatives as usual. If the same problem pops up again it can be wise to take further action.
If it is a disease KNOWN to be inherited, the actions might look a little bit different depending on how it is inherited. Here are some suggestions in how to handle the problem in some different ways. Please note that this is general recommendations. In specific cases there might be a reason to adjust the actions somewhat.
- If the disease is caused by a dominant gene, one has to try and find out which one of the parents carrying it, and if other relatives also might be carrying it. And those who have the disease should not be taken into breeding.
- If the disease is caused by a recessive gene one doesn't of course breed on the sick animal and the parents should also be taken out of breeding, since one knows for sure that they both must be carrying the gene. Possible siblings to the sick kitten shouldn't be sold for breeding, if they are not already sold.
- If the disease is of polygenetic inheritance one should not breed on the sick cat and one shouldn't repeat the same combination that resulted in the afflicted kitten. Be especially observant on littermates and other close relatives, but do not as a rule take them out of breeding, unless they show any signs of problem themselves. If a cat seems to give a high frequency of afflicted cats, even though he/she doesn't have any problems, this cat must of course be taken out of breeding.
Problems in the breed
- Gather all information about the disease! At what age can one see the first signs in an affected cat? How does it manifest itself in the cats? Is it serious? Is there any cure?
- Maybe one can see how it is inherited? Is it a dominant gene, a single recessive gene, sex-linked inheritance, polygenetic?
- Are there ways of testing the cats for the disease, in order to get a quick answer to who is carrying the gene/genes?
- Try and find out approximately what percentage of the population that is affected!
If it is a serious disease, that will affect quite a lot of cats among the breed, it might be useful to make a special health program in order to cope with the problem. How one organizes the health program depends somewhat on the answers to the above mentioned questions. The most important part in all health programs is to spread information. One has to spread information of the symptoms of the disease, development, inheritance, and possible tests available. This can be done through club magazines, brochures, web sites, meetings in breed associations and seminars. Quite often there is resentment among breeders to deal with, or even admit, health problems existing within the breed. This is not often because the breeders don't care if the cats get sick or not, but is often due to fear of the unknown and lack of knowledge. If one doesn't know how to deal with the problems it feels awkward to talk about them. Many are afraid of other breeders' panic reactions, that others will demand whole breeding lines to be neutered if a case of the disease is known. Unfortunately such a fear is not always irrelevant. Due to the same lack of knowledge there are incidents when breeders want to take drastic actions in order to quickly solve the problem within the breed. It is as crazy as putting your head in the sand and pretending there are no problems. There are only two types of panic reaction. And none of them will benefit the cats, of course. The measures taken must be reasonable and in proportion to the difficulty of the problem. Under no circumstances should one select so hard that more than 1/3 of the cats in the population are taken out of breeding due to one and the same health problem. If one takes too drastic measurements the whole breeding population might be too small, and this might cause problems with MORE genetic diseases in the population just because of the diminished breeding population. And this was exactly what we did not want to bring about!
Hence, the foundation of a health program is information, information and yet again information!
Then it might be also be cause for registering cases of disease and possible test results. If one decides to do this, one should also consider the register to be open for all people to take part of the results. This has been a conclusive factor in order for a health program to give good results. Lack of openness will only cause gossip and speculations, while facts will effectively end all discussions of that type. And then all the energy might be put into doing something useful about the problem.
Conclusion
- Healthy cats: Only breed on healthy cats!
- Effective population: Make sure you have a big enough effective population! The inbreeding coefficient on 5 generations should be maximum 2.5% in average on all the matings done. Maximum approximately 5% on 10 generations.
- Inbreeding: Don't make individual matings between cats that are related more closely than cousins (maximum approximately 6.25% on 4-5 generations)!
- Openness: be open about eventual problems in contact with other breeders. It will be helpful to all in the long end!
- No panic: Avoid both hiding your head in the sand and demanding a too severe selection against possible health problems!